生命体如何从一个受精卵发育为完整个体?成体组织在损伤后又如何启动再生修复?Wnt信号通路在这些过程中发挥着核心“开关”作用。这个“开关”究竟如何被启动,是生命科学领域长期关注的重要科学问题。
北京时间5月27日,上海科技大学生命科学与技术学院许文青教授团队在国际学术期刊《细胞》(Cell)在线发表题为“Structural basis of Wnt signalosome extracellular complex assembly”的研究论文,首次解析了高分辨率的Wnt信号体胞外复合物三维结构,系统揭示了天然Wnt配体启动经典Wnt信号通路的分子机制。这一科学谜题的解答,为理解发育、再生和疾病发生提供了关键结构基础,也为结直肠癌、肝癌、骨质疏松、组织纤维化和代谢性疾病等Wnt通路相关重大疾病的治疗,以及再生医学工具开发提供了重要蓝图。
Wnt信号通路在细胞增殖、迁移、分化及凋亡等关键生物学过程中发挥重要作用,广泛参与胚胎发育、组织再生与稳态维持。该通路失调可导致发育缺陷、组织纤维化、代谢紊乱及癌症等多种疾病。其中,经典Wnt/β-catenin信号通路是研究最深入的途径,其启动关键在于Wnt配体、Frizzled (Fzd)受体及LRP5/6共受体组成的Wnt信号体复合物。
围绕经典Wnt信号通路的激活机制,学界长期存在两种主要假说:一种是基于Fzd作为G蛋白偶联受体(GPCR)属性的“变构激活模型”;另一种则认为Wnt作为“交联剂”促使受体聚集,从而触发信号转导,即“聚集激活模型”。然而,由于重组Wnt蛋白制备极其困难、复合物稳定性差且高度异质性,该复合物的结构解析在过去近30年里进展缓慢,成为领域内公认的“硬骨头”。
历时近六年攻关,许文青团队终于攻克了这一难题,成功制备出稳定的Wnt3a/Fzd8/LRP6三元复合物,并利用单颗粒冷冻电镜技术解析了复合物高分辨率三维结构。在此基础上,研究团队提出了“聚集激活”的Wnt信号通路启动模型。
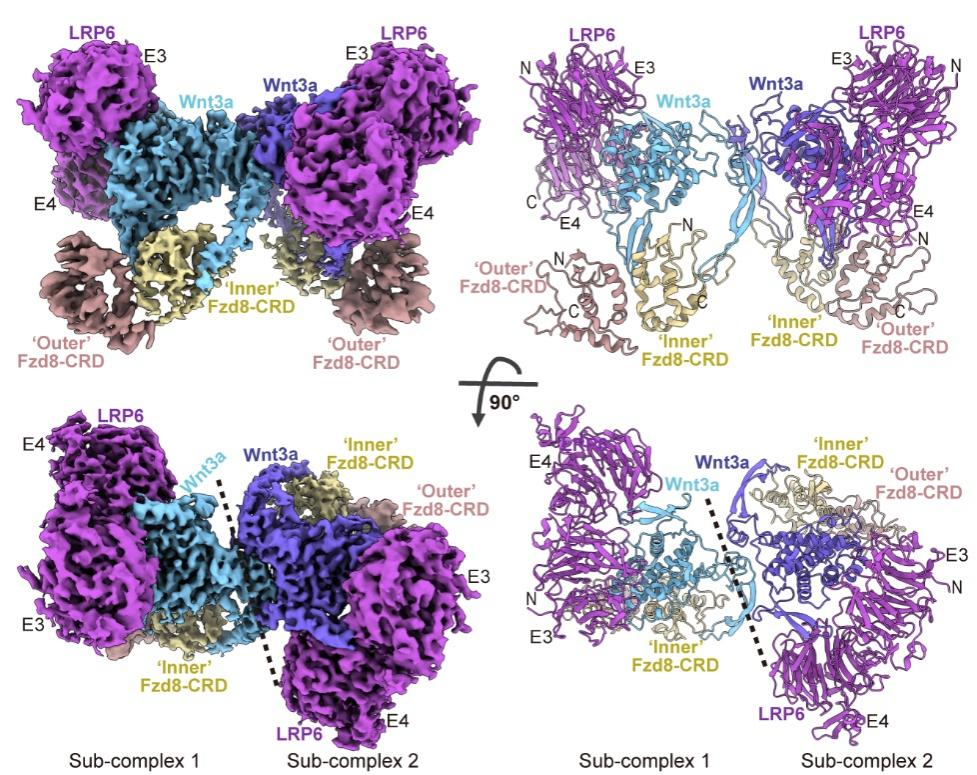
图1.Wnt3a信号体复合物结构。Wnt3a/Fzd8-CRD/LRP6-E3E4三元复合物的冷冻电镜密度图(左图)和结构模型(右图)。
该复合物结构显示,Wnt、Fzd和LRP以2:4:2的化学计量比组装。其中,两个Wnt3a配体形成同源二聚体,构成整个信号体的核心骨架。这对“双生子”各自“牵拉”两个Fzd8受体和一个LRP6共受体,将四个受体和两个共受体“交联”成一个有序的功能单元。这一发现为“聚集激活模型”提供了直接而有力的结构证据。
研究团队进一步通过定点突变、信号活性检测及活细胞成像等多种生化与细胞实验证实,破坏Wnt3a的二聚化界面会完全阻断受体聚集和下游信号激活,凸显了Wnt二聚体在信号体组装中的核心作用。该研究还揭示了信号传导的下一步机制:Wnt3a诱导的Fzd四聚化,可能在细胞内侧创造多价信号蛋白结合平台,例如招募Dvl等下游信号分子,从而协同放大并传递信号。此外,研究团队精准解析了Wnt与LRP6的相互作用界面,并鉴定出具有更高活性的Wnt3a突变体。这些发现不仅加深了人们对天然Wnt信号启动机制的理解,也为人工设计Wnt模拟物、开发Wnt通路调控药物以及构建更高效的类器官培养和组织再生体系提供了结构基础。
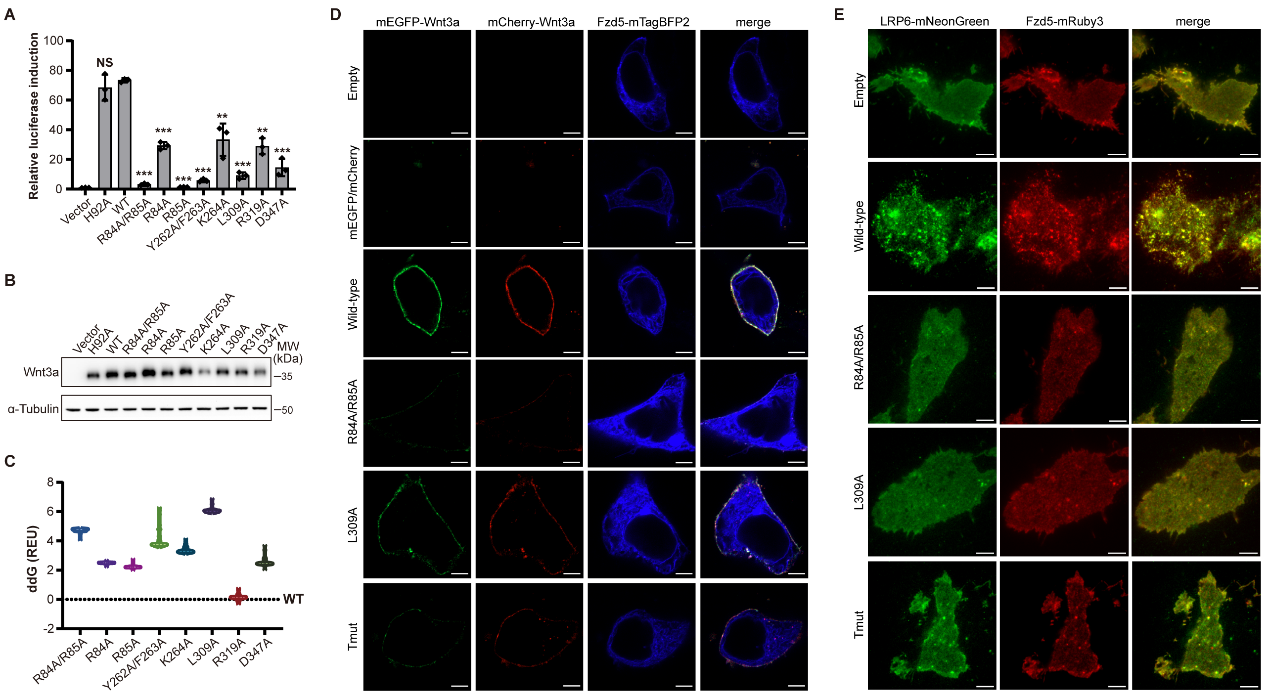
图2. Wnt3a-Wnt3a二聚体在Wnt信号体组装和下游信号传导中发挥关键作用。(A)利用TOPFlash荧光素酶报告实验检测不同Wnt3a突变体的信号活性。(B)通过Western blot检测不同Wnt3a突变体的分泌情况。(C)Wnt3a突变体与野生型蛋白在Wnt3a同源二聚化界面的结合自由能差异(ddG)。(D)ΔFZD1–10 HEK293T细胞mEGFP-Wnt3a/mCherry-Wnt3a的激光共聚焦显微镜成像。(E)不同Wnt3a变体处理后的Fzd5-mRuby3和LRP6-mNeonGreen的全内反射荧光显微镜成像。
本工作首次解析了高分辨率的天然Wnt信号体胞外复合物三维结构,为理解Wnt配体如何同时结合Fzd受体与LRP5/6共受体、形成具有功能活性的Wnt信号体并激活下游通路提供了关键信息,也为Wnt诱导受体聚集激活模型提供了直接结构证据。
值得一提的是,该成果并非孤立突破,而是许文青教授长期深耕Wnt信号通路研究的延续和深化。许文青教授是国际上最早利用结构生物学方法研究Wnt信号通路的科学家之一。他在MIT博士期间的研究就揭示了与Wnt通路共同调控胚胎发育的PAX家族的首个结构(Cell, 1995),在哈佛大学博士后期间解析的首个Src激酶家族结构及其调控机制至今仍是国内外教科书中细胞信号调控的经典内容(Nature,1997)。他1999年入职美国华盛顿大学(西雅图)担任tenure-track助理教授,并于2004、2009年分别成为终身副教授和终身正教授。他的课题组围绕经典Wnt通路的核心调控节点开展了系统研究,先后解析和阐明了β-catenin与TCF/LEF转录因子、Axin等关键调控因子的相互作用机制,揭示了Wnt信号在细胞核内如何驱动靶基因表达,以及Wnt信号在细胞质中如何受到多种蛋白质修饰的调控(Cell, 2000; Nature, 2007, 2015, 2018)。这些系列研究为理解Wnt信号的“输出端”和“调控端”奠定了重要结构生物学和化学基础。
许文青教授于2019年全职加盟上海科技大学。回国后,他将长期积累的Wnt研究进一步推进到信号通路的最上游——即天然Wnt配体如何在细胞表面同时招募Fzd受体和LRP5/6共受体,并形成真正具有信号启动能力的受体复合物。本次发表于Cell的研究,正是Wnt信号通路系统性研究的关键一步:它首次从结构层面回答了经典Wnt/β-catenin信号通路“如何被打开”的核心问题,也使该团队在Wnt通路研究中从细胞内信号转导机制,进一步拓展到细胞膜外信号起始机制,形成了从“信号如何被关闭、如何被读取”到“信号如何被启动”的完整研究链条。
从更长远看,此次突破为后续转化研究打开了新的发展空间。一方面,Wnt/Fzd/LRP三元复合物结构可作为药物研发的重要模板,有助于开发针对Wnt异常激活相关肿瘤、组织纤维化和代谢疾病的抑制剂或抗体药物;另一方面,该结构模型为肺、肝、肠道、心肌等组织再生研究及类器官培养体系优化提供了理性设计的新思路。随着上海科技大学在冷冻电镜、蛋白质设计、合成生物学、细胞成像、化学生物学和转化医学等平台能力的持续建设,许文青团队有望围绕Wnt信号体结构与功能,进一步发展出从基础机制解析到疾病干预和再生医学应用的系统研究方向。
上海科技大学生命科学与技术学院许文青课题组岳丹博士为论文第一作者,许文青教授为论文通讯作者,上海科技大学为论文第一完成单位。
 沪公网安备 31011502006855号
沪公网安备 31011502006855号